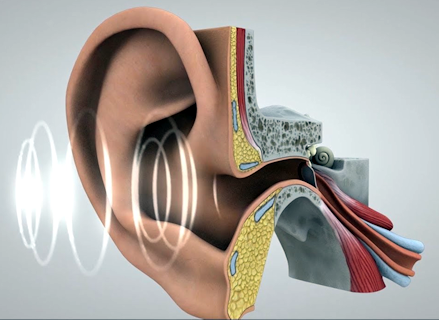
Physiology of Hearing
The pressure pulses that are responsible for the sensation of sound interact with the ear to produce those sensations. The video below illustrates how this interaction occurs. Pressure pulses travel through the air and into the ear canal. At the inner end of the canal, the pulses strike the ear drum. The pulses cause the drum to move in and out. The motions of the ear drum are transferred to a series of three small bones that transmit the motion to the cochlea. In the cochlea the motion is converted into electrical signals that travel along the auditory nerve to the brain. The brain interprets these electrical signals as sound.
Sound waves are first collected from the environment by the pinna (pinnae is the plural), the stiff, convoluted structure on the side of the head that is usually called the ear. The shape of the pinnae play an important role in localizing the source of a sound. Sound waves are directed into the ear canal, which extends about 2.5 cm (1 inch) into the head. The size of this canal enhances frequencies between about 2000 and 6000 Hz. The canal also protects the delicate tympanic membrane, the “ear drum,” at its inner end. The tympanic membrane is a small, thin sheet of skin that moves in and out in response to the pressure changes that form sound waves.
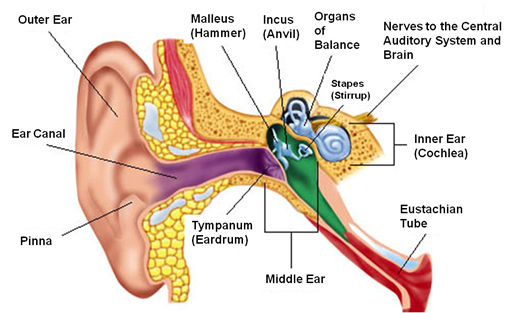
On the inside of the tympanic membrane is the middle ear, which consists of three tiny bones, the ossicles, that amplify sound waves. The first of these bones, the malleus, is connected to the tympanic membrane at one end and to the second ossicle, the incus, at the other. The incus, in turn, is connected to the third ossicle, the stapes, which transmits the vibrations of sound to another membrane, the oval window, which forms the border between the middle ear and inner ear. The names of the bones are based on their shapes or actions. The malleus resembles a mallet or hammer. It pushes on the middle bone, whose name comes from the Latin for anvil, incus. The innermost bone resembles a stirrup, which in Latin is stapes.
The ossicles, the smallest bones in the human body, amplify sound vibrations in two ways. First, the joints between the bones are hinged in a way that makes them work like levers, a modest amount of movement on one side of the joint becomes larger on the other. The lever action increases the pressure change by about a third. Second, the ossicles concentrate the energy from a larger area, the tympanic membrane, to a smaller area, the oval window. The tympanic membrane has an area about 18 times that of the oval window.
The ossicles are also essential in transferring the motion of the tympanic membrane in response to pressure changes in air to motion of a fluid within the inner ear. It takes more energy to move the fluid than to move air. If the fluid of the inner ear were directly behind the tympanic membrane, the membrane would be much less responsive to the pressure variations in sound waves. Sound waves in air would simply be reflected from the membrane rather than cause it to move.
The ossicles also play a role in protecting the inner ear from loud sounds. The middle ear has two muscles attached to the ossicles, one to the malleus and the other to the stapes. These are the smallest muscles in the human body. When a loud sound is detected, these muscles become tense, restricting the motions of the malleus and stapes, thus preventing the energy of further loud sounds from being transmitted to the inner ear. These muscles can protect the ear from sustained loud sounds. However, because there is a slight lag between detecting the loud sound and the tensing of the muscles, they cannot protect against sudden loud sounds, such as an explosion. These muscles also tense when we move our jaw, such as when eating or talking, which keeps our hearing from being overwhelmed by sounds from our own bodies.
Within the inner ear motion generated by sound waves is translated into electrical neural signals. The major structure within the inner ear is the cochlea, a coiled tube whose name is from the Greek for snail, kochlos, since its coiled structure resembles a snail’s shell. The cochlea is about 4 mm in diameter, about the size of a peppercorn. However, if it were uncoiled, its tube would be about 35 mm (almost 1.5 inches) long. The tube is filled with a watery fluid, called the perilymph, and is divided into three parallel canals: the tympanic canal, the vestibular canal, and the middle canal. As its name suggests, the middle canal lies between the other two. The two outer canals are connected to each other at one end by a small opening, called the helicotrema.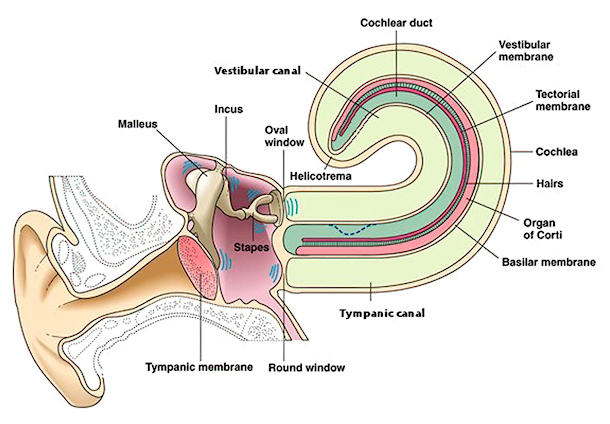
The three canals are separated from each other by two membranes. The vestibular membrane separates the vestibular canal and the middle canal. The basilar membrane separates the middle canal from the tympanic canal. The basilar membrane forms the base of the cochlear partition, a complex structure through which sound waves are converted into neural signals.
Variations in pressure on the tympanic membrane cause it to move in and out. These motions are transmitted by the malleus and incus to the stapes. This causes the stapes to push and pull on the flexible oval window at the base of the cochlea. The movement of the oval window causes pressure changes inside the vestibular canal. An inward push increases the pressure. Because the fluid is nearly incompressible, the increased pressure causes a bulge to form in the vestibular canal (represented by the dashed line in the figure). This bulge travels from the base of the cochlea to its apex. As it travels, its energy is gradually dissipated by the stretching of the cochlear membranes. By the time the bulge has reached the other end of the vestibular canal, its energy is usually spent, and the bulge has disappeared. For particularly loud sounds, however, the pressure may reach the opening at the end and be transmitted back through the tympanic canal. If any should travel the length of the tympanic canal, its remaining energy will be dissipated through a membrane at its end, the round window.
When a bulge occurs in the vestibular canal or tympanic canal, it puts pressure on the middle canal which lies between them. This pressure causes the cochlear partition to move, either down when there is a bulge in the vestibular canal, or up when the tympanic canal bulges. These movements in the cochlear partition are translated into neural signals by structures that run along the top of the basilar membrane. These structures, collectively called the organ of Corti, contains specialized neurons called hair cells. There are two types of hair cells, inner and outer. These cells are arranged in rows along the length of the basilar membrane, one row of inner and three rows of outer hair cells. There are about 3,500 inner hair cells and 12,000 outer hair cells in the organ of Corti. Each of the hair cells has several hairlike projections from the cell membrane, called stereocilia. Over the tops of the stereocilia is a membrane called the tectoral membrane.
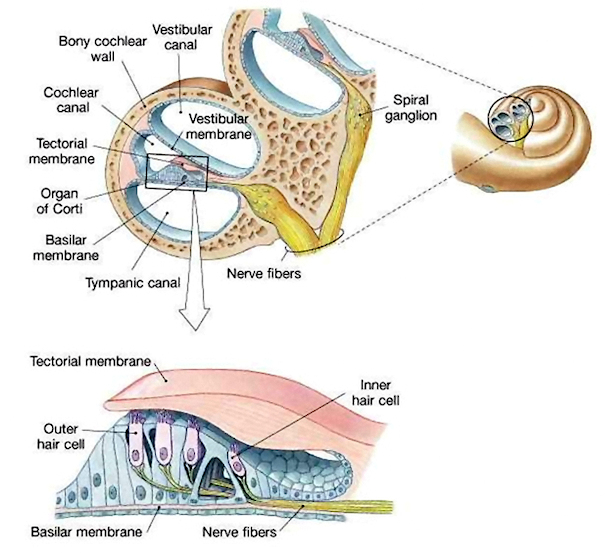
The transformation of sound waves into neural signals begins when the stereocilia bend. The stereocilia bend when a bulge in the vestibular canal passes. The bulge moves the cochlear membrane up. This motion causes the organ of Corti to move up. When the organ of Corti moves up, the stereocilia of the hair cells bend because they rub against the tectorial membrane and because their motion is resisted by the fluid filling the middle canal.
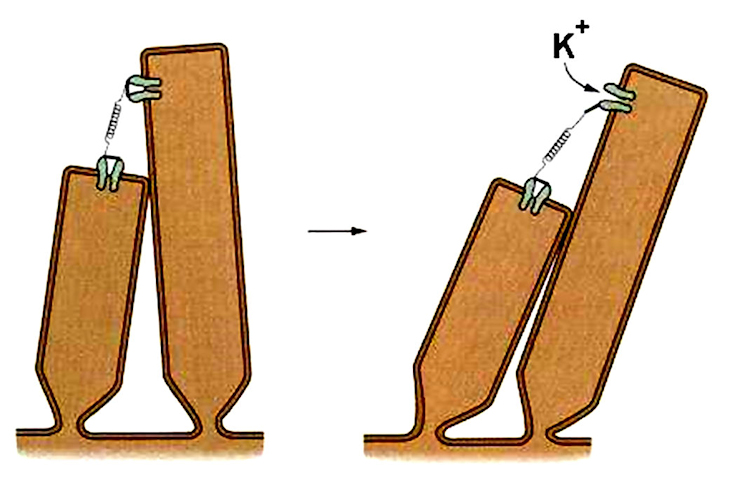
When the stereocilia bend, pores in their membranes open, allowing potassium ions in the surrounding fluid to flow into the cell. This changes the charge inside the stereocilia and changes the electrical potential (voltage) across the membrane of the hair cell. This potential change causes the hair cell to release neurotransmitter molecules from inside, and these neurotranmitters stimulate the firing of auditory nerve fibers that have synapses near the hair cell. The opening of ion pores in hair cells as a result of physical motion is the reason for their short response times and sensitivity. Unlike the case in other senses including vision, depolarization does not wait on a cascade of biochemical reactions.
When the bulge in the vestibular canal passes, the stereocilia move back to their original positions, and the electrical potential dissipates. Hair cells are capable of responding to repeated stimuli in as little as 10 microseconds. This is much faster than the visual receptor cells in the retina, whose response time is hundredths of a second. The hair cells also recover from firing much more rapidly than retinal cells. It takes several seconds for vision to begin functioning when a person enters a darkened theater from a bright, noisy hallway. However, the hushed sounds in the darkened theater can be heard immediately.
The hair cells are also extremely sensitive. The amount of movement in the stereocilia that produces a neural signal is very small. At the threshold of hearing, the stereocilia move about 100 picometers, which is less than the diameter of a carbon atom. Such a motion in the stereocilia can be produced when the eardrum moves by only 10 picometers, less than the diameter of a hydrogen atom. The pressure variations that produce such a movement in the ear drum are barely ten times the random fluctuations in pressure due to movement of air molecules. If our hearing were only a little more sensitive, we could hear a background hiss created by colliding air molecules!
The firing of the auditory nerve fibers is the last step in the conversion of sound waves into neural activity. The overall process begins when an air-pressure wave is directed by the pinna into the auditory canal to the tympanic membrane (eardrum), which oscillates along with the wave. The tympanic membrane moves the malleus, which moves the incus, which moves the stapes, which pushes and pulls on the oval window of the cochlea. Movement of the oval window causes bulges to move down the length of the vestibular canal. These bulges move the middle canal up and down as they pass. This up-down motion causes the tectoral membrane to slide over the hair cells, bending the stereocilia attached to them. The bending of the stereocilia opens pores in the membrane of the hair cells, allowing ions to cross the membrane and changing the cell polarization and releasing neurotransmitters into the synapses between the hair cells and auditory nerve fibers. In response to the neurotransmitters, the auditory nerve fibers carry neural pulses to the brain.
Coding of Amplitude and Frequency in the Cochlea
Two of the fundamental aspects of sound are detected and encoded in the cochlea, namely loudness and pitch. These aspects of perceived sound are manifestations of properties of sound waves. Loudness is determined by the amplitude of a sound wave, and pitch is governed by the frequency of the wave.
As the amplitude of a sound wave increases, the tympanic membrane moves farther in and out with each pressure fluctuation. The resultant bulge in the vestibular canal becomes larger, which causes the cochlear partition to move farther up and down. This causes the hair cells to bend farther back and forth, which causes more neurotransmitter to be released. Increased amounts of neurotransmitter cause the auditory nerve fibers to fire more rapidly. Thus, a sound wave of larger amplitude causes a higher firing rate of the neurons that send information to the brain.
The coding for frequency is more complicated. The basilar membrane which contains the hair cells is not uniform along its length. It actually becomes wider from the oval window to the apex of the cochlea, and it is thicker near the window than at the apex. Through these differences, the basilar membrane is tuned along its length to different frequencies. Higher frequencies bend the narrower, thicker regions of the membrane more, while lower frequencies bend the wider, thinner regions more. Thus, higher frequencies will cause more rapid firing of the auditory neurons near the oval window, while lower frequencies cause more rapid firing of auditory neurons near the apex of the cochlea.
